Plant Cell [2021IF=12.085]-植物所科(kē)研人(rén)員(yuán)揭示可(kě)變剪接調控光(guāng)形态建成的分(fēn)子機(jī)制
植物所科(kē)研人(rén)員(yuán)揭示可(kě)變剪接調控光(guāng)形态建成的分(fēn)子機(jī)制
太陽光(guāng)不僅是植物光(guāng)合作(zuò)用的能量來(lái)源,也是一種重要的環境信号,調節植物的生(shēng)長發育進程。其中幼苗光(guāng)形态建成受光(guāng)質、光(guāng)量的精确調控,涉及不同的光(guāng)受體(tǐ)和一系列信号調控因子。COP1是光(guāng)形态建成的一個明星蛋白(bái),它作(zuò)爲一種E3泛素連接酶,與目标蛋白(bái)互作(zuò)并促進其降解,COP1在植物和動物中廣泛存在。真核生(shēng)物的基因大(dà)多含有内含子,轉錄形成前體(tǐ)mRNA,通過動态剪接組裝,形成成熟的mRNA。可(kě)變剪接也稱選擇性剪接,是一種非常重要的基因轉錄後調控機(jī)制,在特定位點發生(shēng)剪接從(cóng)而生(shēng)成多種mRNA,翻譯成不同蛋白(bái)質,在相(xiàng)應的發育時期發揮重要功能。近年(nián)來(lái),有研究通過轉錄組分(fēn)析了光(guāng)照(zhào)對于可(kě)變剪接的調控現象,但(dàn)對其中的具體(tǐ)調控機(jī)制并不清楚。
中科(kē)院植物所林榮呈研究組鑒定到RNA解旋酶UAP56是調控光(guāng)形态建成的一個新因子。UAP56在拟南(nán)芥中有兩個高度同源的成員(yuán)UAP56a和UAP56b,位于染色體(tǐ)上相(xiàng)鄰的位置。研究人(rén)員(yuán)從(cóng)遺傳學入手,發現UAP56a和UAP56b功能冗餘,但(dàn)兩者純合緻死,利用uap56a純合與uap56b雜合的突變材料,證明UAP56參與到光(guāng)敏色素和隐花色素光(guāng)受體(tǐ)介導的光(guāng)形态建成信号通路(lù)中,是一個負向調控因子。通過尋找互作(zuò)蛋白(bái),發現UAP56與COP1以及剪接輔助因子U2AF65直接相(xiàng)互作(zuò)用。研究人(rén)員(yuán)進一步利用Nanopore三代轉錄組測序技術(shù),發現在黑(hēi)暗及光(guāng)照(zhào)條件(jiàn)下,UAP56和COP1調控了大(dà)量基因的轉錄與可(kě)變剪接,其中包括很多植物生(shēng)長發育、光(guāng)合作(zuò)用以及光(guāng)信号因子等相(xiàng)關基因都(dōu)受二者轉錄或剪接的調控。進一步進行RNA免疫沉澱實驗,顯示UAP56和COP1可(kě)以直接結合到剪接體(tǐ)和靶基因的mRNA上,表明UAP56和COP1可(kě)以通過作(zuò)用于剪接體(tǐ)從(cóng)而直接調控光(guāng)信号介導的選擇性剪接。
該研究工(gōng)作(zuò)一方面鑒定到光(guāng)形态建成的新成員(yuán)UAP56,另一方面發現了COP1作(zuò)爲可(kě)變剪接調節因子的新功能,揭示了UAP56與COP1共同通過調節下遊基因可(kě)變剪接來(lái)調控光(guāng)形态建成的分(fēn)子機(jī)制和調控網絡,增加了對植物光(guāng)信号轉導機(jī)制的認識。同時,UAP56在植物、酵母、動物等都(dōu)廣泛存在,具有保守的RNA解旋酶結構,UAP56-COP1的作(zuò)用方式爲研究其它物種的類似機(jī)制提供了重要參考。
該研究成果于2022年(nián)8月3日(rì)在線發表于國(guó)際學術(shù)期刊The Plant Cell上。植物所已畢業的李洋博士爲第一作(zuò)者,林榮呈研究員(yuán)爲通訊作(zuò)者。該研究得(de)到國(guó)家重點研發計(jì)劃、國(guó)家自(zì)然科(kē)學基金以及中科(kē)院項目的支持。
文章(zhāng)鏈接:https://doi.org/10.1093/plcell/koac235
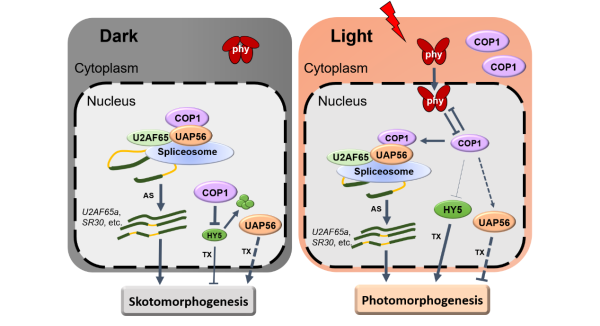
UAP56-COP1與剪接體(tǐ)互作(zuò)調控RNA可(kě)變剪接進而影(yǐng)響光(guāng)暗形态建成的工(gōng)作(zuò)模型